
In natürlichen und wirtschaftlich genutzten Wäldern entstehen laufend die verschiedensten Totholz-Strukturen. Die Biomasse stehender und liegender Baumleichen, von Astmaterial oder von Strünken unterliegt nun einem qualitativen und quantitativen Zerfallsprozess, an dem neben Pilzen und Bakterien auch zahlreiche Wirbellose, insbesondere Insekten, beteiligt sind, welche die Zellulose und andere Pflanzenstoffe verwerten (AUSMUS, 1977). Totholz bietet zudem ein grosses Angebot an verschiedenen Nischen und beherbergt eine artenreiche Arthropodenfauna.
Bereits in der ersten Hälfte des Jahrhunderts wurden ökologisch-faunistische Untersuchungen zu einzelnen Baumarten und deren Zerfallsstadien sowie den damit verbundenen Insektengilden unternommen (KROGERUS, 1927; DERKSEN 1941; SCHIMITSCHEK, 1953a, b; WALLACE, 1953). Die meisten dieser Arbeiten behandeln die Zusammensetzung der Collembolen- und Coleopteren-Fauna auf einzelnen Baumarten. Die Dipteren, speziell die Fliegen (Diptera, Brachycera und Cyclorrhapha), wurden aber nicht oder nur ungenügend abgehandelt. Bis heute besteht ein Mangel an biologischem und faunistischem Wissen über Arten, die mit Totholz in Verbindung stehen. Neuere Artenlisten und ökologische Zusammenstellungen sind nicht vollständig und basieren vor allem auf Kescherfängen oder Zuchtversuchen (vgl. PERRY & STUBBS, 1978).
In den Achtziger- und Neunzigerjahren gewann die Totholz-Forschung wegen der Forderung nach naturnahem Waldbau, Rückführung von Wirtschafts- in Urwälder (BROGGI, 1992), Artenschutz (v.a. bei Käfern) und wegen der Borkenkäfer-Problematik im Zusammenhang mit dem Waldsterben wieder an Aktualität (PFARR, 1990). Durch das Ausräumen und die starke Bewirtschaftung der Wälder sind wohl viele Insektenarten, namentlich einige Käferarten, weitgehend verschwunden (vgl. PFARR, 1990). Über die Fliegenfauna ist auch zu diesem Punkt nichts bekannt.
Das Ziel dieser Arbeit besteht darin, die Fliegenfauna auf Totholz am Beispiel der Wurzelstöcke quantitativ und qualitativ zu erfassen, ein Inventar zu erstellen sowie Angaben zu den Lebensweisen und Phänologien der Fliegen zu machen. Ferner werden verschiedene Baumarten und Abbaustadien auf ihre Artenidentität verglichen sowie die ökologischen Ansprüche der Fliegen an das Totholz abgeklärt. Die zwei angewendeten Methoden (Emergenzfalle und Kescher) werden verglichen und ihre Anwendung im Bezug auf die Totholz-Forschung kurz diskutiert. Naturschutzmassnahmen im Wald zur Förderung und Erhaltung der Dipteren werden zusammengefasst.
Die Ordnung der Diptera ist eine der grössten Insektengruppen überhaupt. Weltweit sind bisher rund 100'000 Arten beschrieben worden. Aus der Schweiz sind mehr als 4000 Arten bekannt (B. MERZ mdl.). Diptera sind euholometabole Insekten und durchlaufen also eine vollkommene Verwandlung von der Larve zur Imago mit einer dazwischen eingelegten Puppenruhe (JACOBS & RENNER, 1988).
Abb. 1: Weibliche Agromyzidae (aus SPENCER,(1976): The Agromyzidae (Diptera) of Fennoscandia and Denmark. Fauna Ent. Scand. 5.1: 15.) als Beispiel einer höheren Fliege. A = Humerus; B = Mesopleura; C = Notopleura; D = Mesonotum; E = Squama; F = Haltere; G = Tergit; H = Ovipositor; I = Sternopleura; J = Coxa; K = Femur; L = Tibia; M = Tarsus.
Die Larve ist vom Imaginalstadium in Aussehen und Lebensweise deutlich verschieden. Dipterenlarven sind normalerweise apod. Nematocera und die meisten niederen Brachycera sind eu- oder hemicephal, d.h. eine sichtbare Kopfkapsel ist entwickelt. Bei den höheren Brachycera (Infraordnung Muscomorpha) ist diese Kopfkapsel reduziert, der Kopf besteht fast nur noch aus den Mundwerkzeugen. Diese apod-acephalen Larven werden als Maden bezeichnet. Die Ernährungs- und Lebensweisen unterscheiden sich oft grundlegend von denen der Imagines. Die Larven leben sowohl aquatisch als auch terrestrisch.
Die Puppe ist bei Nematoceren und niederen Brachyceren bis und mit der Infraordnung Tabanomorpha eine Mumienpuppe (Scheiden der Körperanhänge durch erhärtete Häutungsflüssigkeit an Körper geklebt), bei höheren Brachyceren (Infraordnung Muscomorpha) eine Tönnchenpuppe, d.h. eine freie Puppe liegt in der vorletzten und letzten, dunkel pigmentierten, sklerotisierten Larvenhaut (JACOBS & RENNER, 1988).
Die Imagines (Abb. 1) sind äusserst vielgestaltig und widerspiegeln die Anpassung an die unterschiedlichsten Lebensweisen. Dipteren besiedeln als Imago nur terrestrische Lebensräume.
Die Ordnung der Diptera ist ein Monophylum, welches durch eine gute Synapomorphie begründet ist: Die Reduktion der Hinterflügel zu Schwingkölbchen, den sogenannten Halteren (McALPINE & WOOD, 1989).
Die Ordnung der Dipteren wird in zwei Unterordnungen eingeteilt, die v.a. anhand der Flügeladerung, der Ausbildung der Fühler und der Gestalt der Puppen vorgenommen wird.

Innerhalb der Brachycera können nach McALPINE & WOOD (1989) vier Infraordnungen unterschieden werden:
Einer der wichtigsten Unterschiede zwischen den ersten drei Infraordnungen und Muscomorpha befindet sich am Kopf. Letztere sprengen ihre Puppenhülle durch die Ausstülpung einer Stirnblase, die später zurückgebildet wird und dann die Ptilinalnaht hinterlässt, welche auch noch im Imaginalstadium erkennbar ist (vgl. Abb.2) (JACOBS & RENNER, 1988).

Abb. 2: Frontalansicht des Kopfes einer niederen (Medetera sp., Dolichopodidae) und einer höheren Fliege (Drosophila sp., Drosophilidae).
Totholz entsteht laufend durch Naturereignisse wie Blitzschlag, Windwurf, Rutschungen, Lawinen sowie durch biologische Alterungsprozesse und gehört zur Dynamik des Lebensraumes Wald. Massenvermehrung und Befall mit holzbohrenden Käferarten können ebenfalls Bäume zum Absterben bringen (STEPHEN & DAHLSTEN, 1976). Daneben spielen auch Pilze, welche noch lebende Bäume durchwachsen, eine wichtige Rolle beim Absterbeprozess (COOKE & RAYNER, 1984). Dies führt zu liegenden Baumleichen oder stehendem Totholz.
Bei der Bewirtschaftung des Waldes durch den Menschen entstehen ebenfalls beträchtliche Mengen an Abfallholz wie Wurzelstöcke, Astmaterial beim Entasten, Reisighaufen, Lagerholz usw. (PFARR, 1990).
Das stehende und liegende tote Holz unterliegt einer Zerfalls-Sukzession, in welcher zahlreiche biotische Faktoren wie Pilze und Arthropoden und abiotische Faktoren den Abbau beeinflussen (Tab. 1).
Das Zusammenwirken von verschiedenen abiotischen Standortsfaktoren wie Gross-, Standort- und Mikroklima mit den verschiedenen Temperatur-Feuchtigkeitskombinationen, hemmt oder fördert den chemischen Abbau und die Lebensprozesse der beteiligten Organismen und bestimmt somit auch die Dauer des Zerfalls. Relief, Länge der Vegetationsperiode und Schneebedeckung sowie die Sonneneinstrahlung haben ebenfalls einen Einfluss auf den Abbau des Holzes (SCHIMITSCHEK, 1953a).
Tab. 1: Abiotische und biotische Faktoren, welche den Zerfall des Totholzes
bewirken (nach SCHIMITSCHEK, 1953a)
ABIOTISCHE FAKTOREN BIOTISCHE FAKTOREN
Klima Niederschlag Arthropoden Coleoptera
Luftfeuchte Diptera
Sonneneinstrahlung Collembola
Schneebedeckung Pilze Weissfäule
Temperatur Rotfäule
Relief Exposition Bakterien
Neigung
Zerfallende Totholz-Strukturen sind interessante biologische Einheiten und wurden von FOG (1977) als Mikroökosysteme klassiert. Zu den Hauptkomponenten eines "Totholz-Systems" gehören (a) das Angebot an totem organischen Material, welches über die Zeit langsam abnimmt, (b) Primärdestruenten (hauptsächlich Basidiomyceten), (c) eine begleitende Mikroflora, die grösstenteils auf die Oberfläche des Holzes beschränkt ist, und (d) eine Wirbellosen-Fauna, welche sich hauptsächlich von der Mikroflora ernährt. Der Zerfallsverlauf hängt von den Interaktionen der einzelnen Gruppen untereinander ab (FOG, 1977).
Zwischen Pilzen und Insekten besteht eine enge mutualistische Interaktion (COOKE & RAYNER, 1984). Holzbohrende Käferarten (v.a. Scolytidae) öffnen das Totholz für Pilze und sorgen für deren weitere Verbreitung, indem sie Sporen in speziellen Organen oder auf ihrem Körper in Bohrgänge eintragen. Diese Saprophyten, v.a. Ambrosia-Pilze, bilden an den Wänden der Käfergänge ein Mycel aus, schliessen die Zellulose enzymatisch auf und machen auf diese Weise das Holz für die Insekten verdaulich. Die Pilze dienen auch als direkte Nahrungsquelle und liefern den Käfern Aminosäuren, Vitamine, Sterole sowie Enzyme, damit diese selber Zellulose aufschliessen können. Das Mycel profitiert dafür von den stickstoffhaltigen Käferexkrementen (COOKE & RAYNER, 1984). Zur Nahrungsaufnahme nehmen xylophage Insekten Holz auf, verdauen einen Teil davon und scheiden den Rest mit anderen Abfallstoffen aus. Dieses feinkörnige Material hat eine vergrösserte Oberfläche und kann mehr Wasser zurückhalten, fördert also das Wachstum der Pilze (FAGER, 1968).
Besiedlung, Verbreitung und Aktivität der Pilze auf Totholz wird durch den Wassergehalt und die Durchlüftung des Totholzes gesteuert. Der Feuchtegehalt muss mehr als 24% des Trockengewichtes des betreffenden Holzteiles betragen. Die Dichte von Destruenten (Pilze, Bakterien), Mikro- und Makroarthropoden nimmt zu, wenn der Feuchtigkeitsgehalt des abgestorbenen Stammes eine gewissen Grad erreicht. Der luftgefüllte Porenraum im Holz darf aber nicht kleiner als 10-20% werden (BASSET, 1986). Ist der Wassergehalt zu hoch, kann der Stamm nicht abgebaut werden (z.B. untergetauchtes Holz) (COOKE & RAYNER, 1984).
Absterbendes Holz ändert im Verlauf der Verwitterung seine strukturellen und chemischen Eigenschaften, durchläuft also verschiedene Abbaustadien, bis es schliesslich zur organischen Substanz des Bodens geworden ist.
A B B I L D U N G (in Arbeit) Abb. 3: Altersklassifikation von stehendem und liegendem Totholz nach MASER et al. (1979).
Im Laufe der Totholz-Forschung haben diverse Autoren Klassifikationen für Abbaustadien von Baumstämmen vorgeschlagen. SCHIMITSCHEK (1953a, 1954) beobachtete die Befallsfolgen der einzelnen Baumarten an Kambialzone und anhand des Zerfalls von Lagerholz und teilte jeder Phase eine charakteristische Insektengesellschaft zu. BRAUNS (1991) und FOG (1977) nutzten das Wissen um den Schlagzeitpunkt der Bäume und zeigten graphisch den Zerfall der einzelnen Strünke nach der Holzernte. Dieses Verfahren ist aber problematisch und stimmt nicht mit dem natürlichen Zerfall überein (schnellerer Zerfall an luftfeuchten, schattigen Standorten als an sonnenexponierten Stellen). MASER et al. (1979) erarbeiteten eine Methode, um die Altersstadien im Feld zu bestimmen (vgl. Abb. 3).
Insekten durchlaufen, wie die holzzerstörenden Pilze, eine qualitative und quantitative Sukzession (DERKSEN, 1941; WALLACE, 1953).
Für die Erstbesiedelung durch Tiere ist der Feuchtigkeitsgehalt des Totholzes ausschlaggebend, welcher von Standortsverhältnissen, Relief und geographischer Lage abhängig ist (SCHIMITSCHEK, 1953a). Im weiteren Verlauf der Zersetzung ändert sich die Insektengilde bedingt durch die verschiedenen Zerfallsstadien. WALLACE (1953) fand v.a. eine grosse Veränderung der prozentualen Anteile der verschiedenen Ernährungstypen (vgl. Abb. 4).
Abbildung (in Arbeit) Abb. 4: Prozentualer Anteil der verschiedenen Ernährungstypen in Föhrenstrünken in verschiedenen Phasen des Zerfalls nach WALLACE(1953).
Dipteren wurden im bisherigen Verlauf der Totholz-Forschung nur ungenügend abgehandelt. Die Angaben zu Arten sind oft nur lückenhaft (z. B. BRAUNS, 1991; PERRY & STUBBS, 1978; SCHMITSCHEK, 1953b; WALLACE, 1953). Neuere Untersuchungen zeigen, dass Dipteren einen grossen Anteil an der Arthropodenfauna von Totholz ausmachen können und deshalb eine nicht zu unterschätzenden Einfluss auf die ganze Biozönose des Totholzes haben (vgl. PFARR, 1990).
Innerhalb einer Totholz-Struktur nehmen die Dipteren bzw. deren Larven, bedingt durch ihre Ernährungsweisen und das Zerfallsstadium des Holzes, ganz bestimmte Nischen ein. Im folgenden soll an wenigen Beispielen die wichtigsten Mikrohabitate und Ernährungstypen aufgezeigt werden. Einige Arten scheinen auch an bestimmte Baumarten respektive an Laub- oder Nadelholz gebunden zu sein.
a) carnivor - parasitoide Lebensweise
Etliche Arten aus den Familien Empididae und Hybotidae, deren
Larven sich räuberisch von anderen Insektenlarven ernähren,
leben unter der Rinde. Vertreter der Gattung Medetera (Dolichopodidae)
leben als Parasitoiden auf Käferlarven in Käfergängen.
Weitere Beispiele: Lonchaeidae, Rhinophoridae
b) fungivore Lebensweise
Wie bereits angedeutet zählen Pilze zu den wichtigsten holzabbauenden
Lebewesen. Dieses Nahrungsangebot nutzen in zahlreiche Vertreter
der Nematoceren, bei den Fliegen sind es in erster Linie Drosophilidae,
deren Larven Pilze und Bakterien im modernden Holz oder unter
der Baumrinde fressen (BÄCHLI & BURLA, 1985).
Weitere Beispiele: Heleomyzidae (Gattung Suillia)
c) xylophag - saprophage Lebensweise
Durch den Zerfall des Holzes verändert sich auch dessen Struktur.
Stark zerfallenes Totholz dient als sogenannter Holzdetritus v.a.
Tipulidae oder gewissen Muscidae als Nahrungsgrundlage (Moderfresser).
Die Larven leben unter Moospolstern (z.B. Tipulidae), unter der
Rinde (Muscidae) und in faulenden Baumhöhlen oder Astlöchern
(z.B. Syrphidae). Muscidae können auch Hohlräume unter
der Rinde besiedeln.
Weitere Beispiele: Rhagionidae, Lonchaeidae
d) Saftflüsse von frisch gefällten Bäumen
Einige Dipteren-Gruppen nehmen Baumsäfte zu sich, wenn z.B.
durch Windwurf oder durch die Holzernte frische offene Kernholzflächen
entstehen. Wichtige Exponenten finden sich bei den Syrphidae und
den Drosophilidae. Saftflüsse dienen in vielen Fällen
nur den Imagines als Futterquelle.
Weitere Beispiele: Periscelidae, Fanniidae
In Totholz leben die Larven von mehr als 200 Dipterenarten aus über 40 Familien (PERRY & STUBBS, 1978). Da aber systematische Untersuchungen über die Dipterenfauna, v.a. über die Fliegen, auf Totholz weitgehend fehlen, beruht die unten aufgeführte Zusammenstellung auf den spärlichen Angaben von BRAUNS (1991), PERRY & STUBBS (1978) und TESKEY (1976).
Die ganze Familien-Systematik folgt vereinfacht den Vorschlägen von McALPINE & WOOD (1989). Die Hybotidae werden als eigene Familie aufgeführt (CHVALA, 1983). Diese Anordnung wird in der ganzen Arbeit beibehalten.
Unterordnung BRACHYCERA
Infraordnung Xylophagomorpha
Familie Xylophagidae
Infraordnung Stratiomyomorpha
Familie Xylomyiidae
Familie Stratiomyidae
Infraordnung Tabanomorpha
Familie Rhagionidae
Infraordnung Muscomorpha
Überfamilie Asiloidea
Familie Therevidae
Familie Scenopinidae
Familie Asilidae
Überfamilie Empidoidea
Familie Empididae
Familie Hybotidae
Familie Dolichopodidae
Sektion Aschiza
Überfamilie Platypezoidea
Familie Platypezidae
Familie Phoridae
Überfamilie Syrphoidea
Familie Syrphidae
Sektion Schizophora
Untersektion Acalyptrata
Überfamilie Nerioidea
Familie Micropezidae
Überfamilie Diopsoidea
Familie Tanypezidae
Familie Psilidae
Familie Megamerinidae
Familie Chyromyidae
Überfamilie Tephritoidea
Familie Lonchaeidae
Familie Otitidae
Familie Pallopteridae
Überfamilie Lauxanioidea
Familie Lauxaniidae
Überfamilie Opomyzoidea
Familie Clusiidae
Familie Acartophthalmidae
Überfamilie Agromyzoidea
Familie Odiniidae
Überfamilie Asteioidea
Familie Periscelidae
Familie Asteiidae
Überfamilie Carnoidea
Familie Milichiidae
Familie Chloropidae
Überfamilie Ephydroidea
Familie Drosophilidae
Untersektion Calyptrata
Überfamilie Muscoidea
Familie Anthomyiidae
Familie Fanniidae
Familie Muscidae
Überfamilie Oestroidea
Familie Sarcophagidae
Familie Rhinophoridae
Familie Tachinidae
Die gesamte Untersuchung wurde im Sihlwald, Kanton Zürich, in der Nähe der Station Sihlbrugg auf 560 - 710 m.ü.M im Bereich zwischen dem Eschentobel (Koordinaten 685/232) und Wüesttobel (684/233) durchgeführt.
Der Sihlwald liegt südlich der Stadt Zürich ca. 15 km Luftlinie entfernt im mittleren Sihltal zwischen der Albis- und der Zimmerbergkette, nimmt eine Fläche von rund 10 km2 ein und ist eines der grössten zusammenhängenden Waldgebiete des schweizerischen Mittellandes (BSU, 1988). Die natürlichen Grenzen des Gebietes werden im Osten durch die Sihl, im Westen durch den Albisgrat gebildet. Die höchsten Punkte liegen auf 914 m.ü.M (Bürglen), die tiefsten auf 480 m.ü.M. Der grösste Teil des Waldes stockt auf der NE-exponierten linken Seite des Sihltals. Kleinere Teile davon nehmen eine SW-exponierte Lage am rechten Sihlufer und am Albis-Südhang ein (vgl. Abb. 6 nächste Seite).
Das Klima der Region ist ein typisches Mittellandklima von ozeanischer Prägung mit einem relativ ausgeglichenen Temperaturverlauf und einem mittleren jährlichen Niederschlag von 1258 mm im nahegelegenen Horgen (Abb. 5). Die mittlere Januartemperatur beträgt -0.7 ¡C, die mittlere Julitemperatur 17 ¡C. Durch die grössere Nähe zur Innerschweiz ist der Einfluss des Föhns bereits spürbar, die durchschnittswerte der Temperatur sind etwas grösser als in Zürich.

Abb. 5.: Klimadiagramme von Zug und Horgen (verändert nach WALTER & LIETH, 1967) A B B I L D U N G (in Arbeit) Abb. 6: Übersichtskarte über den Sihlwald und Umgebung (Kartenausschnitt aus der Landeskarte der Schweiz 1:50'000, Blatt 225 Zürich). Die Lage des Untersuchungsgebietes ist rot markiert.
Zwischen dem 13. April und dem 25. Juli wurden an insgesamt 16 Tagen (in Tab. 2 schwarz eingezeichnet) aus den Fallen und in den unter Kapitel 3.2.3. bezeichneten Flächen und Strukturen Fliegen gesammelt. An acht Tagen wurden nur die Emergenzfallen bearbeitet (dunkelgrau), an einem Tag (25.7.) wurde nur mit dem Streifnetz gesammelt (hellgrau) (vgl. Tab. 2). Somit wurden also an 25 Tagen Insekten gefangen.
Tab. 2: Übersicht über die Sammelaktivität während der Feldsaison vom 19.4.-7.7.94 im Sihlwald (Erklärungen s. Text).

Ein kompletter Sammeltag setzte sich zusammen aus dem Entleeren der Emergenzfallen am Morgen zwischen 9.00 Uhr und 12.30 Uhr und dem Handfang mit dem Streifnetz am frühen Nachmittag zwischen 12.00 und 15.00 Uhr. Der Zeitaufwand richtete sich nach der Menge der vorhandenen Fliegen.
Für die Bezeichnung der Fallen werden in der ganzen Arbeit die gleichen Abkürzungen benutzt und sind an dieser Stelle kurz erklärt. Die Standorte der Fallen sind in Abb. 8 eingetragen.
1. Buchstabe A = Schlagzeitpunkt 81/82
B = Schlagzeitpunkt 88/89
C = 91/92 resp. 92/93
2. Buchstabe B = Buche
F = Fichte
A = Ahorn
Zahl 1 = erste Falle bei entspr. Baumart und Altersstufe
2 = zweite Falle bei entspr. Baumart und Altersstufe
Für diesen Teil der Untersuchung wurden 14 Baumstrünke (=Stubben) von drei Baumarten (Bergahorn (Acer pseudoplatanus), Buche (Fagus sylvatica) und Fichte (Picea excelsa )) (Nomenklatur für sämtliche Pflanzen folgt LANDOLT et al. (1986)) in drei verschiedenen Altersstufen (2, 5 bzw. >10 Jahre) nach der Holzernte einbezogen (vgl. Tab. 3). Das Strunkalter und die für diese Untersuchung geeigneten Flächen sowie die Strünke wurden anhand von Schlagkarten des Stadtforstamtes Zürich und während einem Rekognoszierungsrundgang Ende März in den Forstabteilungen 4 bis 10 des Sihlwaldes ermittelt. Bei der Auswahl der Strünke wurde darauf geachtet, dass sie von vergleichbarer Grösse waren (d.h. Stammdurchmesser und Höhe). Innerhalb der ausgewählten Fläche sollte das Gelände nach Möglichkeit eine ähnliche Neigung und Exposition aufweisen.
Tab. 3: Aufteilung der Emergenzfallen auf die einzelnen Baumarten und Altersstadien Baumart/Altersstadium Bergahorn Fichte Buche TOTAL A (83/84) - 2 2 4 B (88/89) 1 2 2 5 C (92/93) 1 2 2 5 TOTAL 2 6 6 14
Die Fallen waren rund und wiesen einen Durchmesser von 90 cm und eine Höhe von 80 cm auf. Das Material bestand aus einer derben Gaze mit 1 mm Maschenweite. Ein seitlich angebrachter Reissverschluss von 35 cm Länge ermöglichte das Absammeln der Tiere innerhalb der Fallen. Beim Aufstellen wurde darauf geachtet, dass sich die Öffnung an der lichtabgewandten Seite befand. Das Gazezelt wurde von Innen durch drei sternförmig verbundene, rechtwinklig gebogene Eisenstangen gestützt. Die Seitenränder der Fallen wurden nach Möglichkeit in den Waldboden eingesenkt und mit Erde so gut wie möglich abgedichtet, was v.a. aber an Hanglagen schwierig war (Abb. 7).
Die Fallen wurden am 13. April 1994 aufgebaut und am 7. Juli 1994 abgebrochen. Dazwischen wurden sie in regelmässigen zeitlichen Abständen (vgl. Tab. 2) kontrolliert. An den Käfigen wurde kein Sammelgefäss angebracht, die darin vorhandenen Dipteren wurden mit Hilfe eines kleinen Streifnetzes von 12x10 cm Seitenlänge und 20 cm Sacktiefe lebend abgefangen und mit Hilfe eines Exhaustors in Gläschen abgefüllt. Der Zeitaufwand für das Leeren aller Zelte betrug rund 3 Stunden. Fallen mit gutem Fangertrag beanspruchten ca. 15-20 min. der Zeit, leere Fangkäfige oder solche mit Einzelindividuen nur rund 5 min. Bei schlechter, nasser Witterung konnte nicht aus den Fallen gefangen werden, da v.a. kleinere Tiere an der Wand der Falle klebten und nicht unbeschädigt entfernt werden konnten.
A B B I L D U N G (in Arbeit) Abb. 7: Fallen B/B1, B/B2 und B/A1 Ende April, Sihlwald bei Sihlbrugg, 690 m.ü.M (Foto: P. Rüegg)
Um sich ein Bild von den verschiedenen Standortsverhältnissen und dem Aussehen der Baumstrünke machen zu können, sind hier alle Fallenstandorte und Wurzelstöcke beschrieben. Die Feldaufnahmen wurden am 25.7. und 16.8. gemacht Die Texte enthalten Angaben zu Flurnamen, Topographie und Waldgesellschaften (nach SCHMIDER et al., 1993). Tabelle 4 fasst die wichtigsten Strunkparameter zusammen.
Die Abkürzungen für die Fallenbezeichnungen sind zu Beginn von Kap. 3.2.2. erklärt.
a) Schönboden
Exposition: NE Höhe über Meer: 610 - 630 m
Bestand: Buchenstrünke in typischem Waldmeister-Buchenwald
(Galio odorati-Fagetum typicum) Fichtenstrünke in Seggen-Bacheschenwald
mit Equisetum maximum (Carici remotae-Fraxinetum Equisetosum maximi).
4 Fallen: A/B1 A/B2 A/F1 A/F2. Schlagzeitpunkt: 81/82
Bemerkung: Die Strünke waren sehr inhomogen. Die Buchenstöcke
waren bereits so zerfallen, dass das Holz einen säulenartigen
Aufbau zeigte und teilweise in die Bodensubstanz übergegangen
war. Die Fichtenstrünke hingegen wiesen äusserlich kaum
Spuren des Zerfalls auf (vgl. Tab. 4). Die Standorte der beiden
Baumarten unterschieden sich deutlich.
b) Egliboden (Buchenfallen)
Exposition: NE Höhe über Meer: 690-710 m
Bestand: Waldhirsen-Buchenwald mit Lungenkraut, Ausbildung mit
Stachys sylvatica (Milio-Fagetum pulmonarietosum, Stachys-Variante
).
3 Fallen: B/B1, B/B2, B/A1. Schlagzeitpunkt: 88/89
Bemerkung: Der Wald war ziemlich stark aufgelichtet. Der Jungwuchs,
der v.a. aus Eschen Fraxinus excelsior bestand, war noch niedrig,
so dass die Besonnung der Strünke z.T. recht stark war. Die
Strünke, insbesondere die Buchen, zeigten kaum Anzeichen
des Zerfalls.
c) Egliboden (Fichtenfallen)
Exposition: NE Höhe über Meer: 660-670 m
Bestand: Waldhirsen-Buchenwald mit Pulmonaria sp. (Milio-Fagetum
pulmonarietosum).
2 Fallen: B/F1 B/F2. Schlagzeitpunkt: 88/89
Bemerkung: Beide Fallen standen in einem relativ dichten Buchen-Jungwuchs,
so dass die Besonnung schwach war. Der Standort war deshalb relativ
feucht und trocknete nach Regengüssen nur sehr langsam ab.
Der Strunk B/F2 zeigte in der Mitte der Stammscheibe ein tiefes,
herausgefaultes Loch, was ein deutliches Indiz für den starken
innerlichen Zerfall war. B/F1 hingegen war äusserlich wenig
zersetzt.
d) Wüesttobel
Exposition: NE Höhe über Meer: 670 m
Bestand: Zahnwurz-Buchenwald mit Allium ursinum (Cardamino-Fagetum
allietosum).
1 Falle: C/A2. Schlagzeitpunkt: 92/93<
Bemerkung: Dieser Strunk
wies keine äusseren Anzeichen des Zerfalls auf. Besondere
Merkmale fehlten weitgehend.
e) Eschentobel
Exposition: SE Höhe über Meer: 560-580 m.
Bestand: Typischer Waldmeister-Buchenwald (Galio odorati-Fagetum
typicum ).
4 Fallen: C/B1 C/B2 C/F1 C/F2. Schlagzeitpunkt: 91/92
Bemerkung: Die Fichten-Stubben schienen grösstenteils intakt
zu sein. Die Buchen waren optisch bereits stärker zerfallen.
Die äussere Erscheinung sprach für einen stärkeren
Abbau als bei B/B1 und B/B2.
Wichtige Mikrostrukturelemente sind Moosbewuchs auf Stammscheibe und Rinde, Pilzfruchtkörper, faulende Löcher im Holz und Besatz durch Pflanzen. Eine grosse Bedeutung kommt dem Zustand der Rinde zu. Abblätternde Rinde kann einer grossen Anzahl von raptorischen Larven Unterschlupf bieten, fest angewachsene hingegen nicht. Als weiterer wichtiger Punkt wurde die Besonnung der Strünke aufgenommen. Starke Einstrahlung führt zu einer Austrocknung des Holzes und damit zu einer Trockensterilisation (BASSET, 1986).
In Tab. 4 wurde die Anzahl Pflanzen auf den Strünken in eine Skala, die von 1-3 reicht, eingeteilt. (1: <=5 Ind. auf Strunk; 2: <=20 Ind/Strunk; 3: >20 Ind./Strunk). Die häufigsten Arten auf den Stubben waren Rubus sp., Oxalis acetosella, und diverse Baumkeimlinge (Fraxinus excelsior, Picea excelsa). Die Zentimeterangaben sind Durchschnitte aus fünf Messwerten. Alle Prozentzahlen wurden geschätzt.
Tab. 4: Weitere Parameter der mit den Fallen untersuchten Strünke im Sihlwald (weitere Erklärungen im Text)
| Falle | weitere Merkmale | |||||||||
| A/B1 | teilweise amorph | |||||||||
| A/B2 | teilweise amorph | |||||||||
| A/F1 | Strunk intakt | |||||||||
| A/F2 | Strunk intakt | |||||||||
| B/A1 | Stammscheibe hart | |||||||||
| B/B1 | Stammscheibe hart | |||||||||
| B/B2 | Stockausschlag | |||||||||
| B/F1 | ||||||||||
| B/F2 | sehr tiefes Faulloch | |||||||||
| C/A2 | unregelm. Stamm | |||||||||
| C/B1 | ||||||||||
| C/B2 | Stockausschlag | |||||||||
| C/F1 | ||||||||||
| C/F2 |
Um den qualitativen Aspekt der Arbeit zu vertiefen, wurde als zweite Sammelmethode der Kescher (=Streifnetz) eingesetzt. Damit konnten weitere Tiere auf grösseren Totholz-Strukturen wie Asthaufen, Stammteile oder Lager- und Langhölzer gesammelt werden.
Der Kescher bestand aus einem kreisrunden Bügel von 50 cm Durchmesser und einem Sack von 70 cm Tiefe aus einer stabilen Gaze (Maschenweite 0.1 mm). An den Bügel wurde ein Holzstiel von 50 cm Länge und 2 cm Durchmesser angeschraubt.
Auf drei totholzreichen Flächen (F1, F2, F3) und sieben speziellen Totholzstrukturen (S1-S7), wurde regelmässig gekeschert. In den Flächen wurden 20-25 min. alle möglichen Totholzstrukturen abgesucht. Für das Fangen über den Strukturen betrug der Zeitaufwand durchschnittlich 7 min. (maximal 12 min.). Der Zeitaufwand richtete sich nach dem Aufkommen an Fliegen. Bei schlechter Witterung oder nasser Vegetation wurde nicht gesammelt. Die Lagen der Flächen und Strukturen sind aus Abb. 8 ersichtlich.
In Anlehnung an die Methode von JANETSCH (1982) wurde das Streifnetz möglichst dicht über das Holz in engen Achterschlaufen gezogen , bis die ganze Oberfläche der betreffenden Struktur abgedeckt war. Die gefangenen kleineren Dipteren wurden dann mit Hilfe des Exhaustors in Sammelgläschen eingesogen. Grössere Tiere wurden einzeln aus dem Kescher gefangen.
Auf einer Vorexkursion am 19. April 1994 wurden die Flächen und Strukturen nach folgenden Kriterien ausgeschieden:
Für die Flächen galt, dass (a) auf einem möglichst homogenen, ungefähr gleich grossen Waldstück mehr oder weniger flächendeckend oder über die ganze Fläche verstreut genügend Totholz vorhanden sein musste und (b) sich die Flächen möglichst durch verschiedenartige Strukturen (Material von Windwürfen, Durchforstungen etc.) unterschieden.
Die Strukturen wurden so ausgewählt, dass (a) kein Zusammenhang zu anderen Strukturen bestand und (b) dass sie eine gewisse Fläche aufwiesen. Dazu eigneten sich v.a. Langhölzer von über 30 cm Durchmesser und einer Länge von minimal 5 m. Asthaufen sollten möglichst wenig, höchstens kurze Stammteile beinhalten, die einen Durchmesser von weniger als 30 cm aufweisen sowie eine Fläche von rund 5 m2 einnehmen. Kleinere Totholzstrukturen wie einzelne Äste usw. wurden für diesen Teil der Untersuchung nicht berücksichtigt.
Die Daten für diese Beschreibung wurden am 16.8. erhoben. Dabei wurden v.a. Bemerkungen zu Strukturen und Baumarten des liegenden Holzes aufgezeichnet. Exposition und Neigung sowie die wichtigsten Pflanzenarten der Flächen wurden ebenfalls im Felde bestimmt. Die Zuteilung der Waldgesellschaften richtet sich nach SCHMIDER et al. (1993). Tabelle 5 fasst weitere Parameter zusammen.
Tab. 5: Parameter zur Beschreibung der besammelten Flächen im Sihlwald ST=Stammteile; AM=Astmaterial; R=Reisig; S=Strünke; + = vereinzelt 123456789012
a) Fläche 1 (F1)
Die Fläche erstreckte sich auf beide Seiten eines kleinen, wenig tiefen Bachtobels im Bereich des Kellerbodens. Die Hänge wiesen eine Neigung von ca. 10 Grad auf und waren SW- resp. NE- exponiert.
In diesem Waldteil lag relativ wenig Totholz, die meisten Strukturen waren anthropogen bedingt (Strünke, Holzhaufen). Eine grosse windgeworfene Buche und eine Esche waren die markantesten Totholzelemente in der Fläche. Die Strukturarmut wirkte sich auch auf den Fangertrag aus, so dass hier meist weniger Zeit gebraucht wurde, um die Strukturen flächendeckend abzusuchen (15-20 min.).
Die Tobelseiten waren durch einen Aronstab-Buchenwald (Aro-Fagetum ) bestockt, höher gelegene Teile durch einen typischen Waldhirsen-Buchenwald (Milio-Fagetum typicum).
b) Fläche 2 (F2)
Dieses Waldstück lag fast eben (Neigung 5 Grad) in NE-licher Exposition zwischen dem Mittleren und Oberen Hebeisenbach. Die Fläche wies ein gutes Angebot an natürlichem, liegenden Totholz auf. Zahlreiche hohe Bäume (v.a. Fichten und Eschen) waren relativ frisch durch den Wind umgeworfen, weitere ältere Baumleichen waren teilweise stark vermodert. In der so entstandenen Lichtung kam bereits viel Eschen-Jungwuchs auf, was das Fangen z.T. erschwerte.
Die verschiedenen Pflanzengesellschaften sind auf dieser Fläche mosaikartig verzahnt. In Wegnähe konnte ein Waldhirsen-Buchenwald, Ausprägung mit Stachys sylvatica (Milio-Fagetum pulmonarietosum ) festgestellt werden, in den übrigen Teilen ein Seggen-Bacheschenwald (Carici remotae-Fraxinetum typicum ) mit Cornus sanguineum resp. mit Allium ursinum.
c) Fläche 3 (F3)
Die dritte Untersuchungsfläche befand sich im Bereich des Wüesttobels und war mit 5-grädiger Neigung nach NE exponiert. Dieser Waldteil wies einen grossen Reichtum an verschiedenem Totholz auf, welches vollständig von der letzten Durchforstung der Fläche im Winter 1992/93 stammt. Im oberen Teil wurden v.a. Strünke, Stammteile und dickes Astmaterial abgesucht. Im unteren Teil war der Waldboden flächendeckend knöcheltief mit Reisig, Rinde und dünnerem Astmaterial übersäht. Einzelne Strünke ragten daraus hervor.
Die Waldgesellschaft war ein Zahnwurz-Buchenwald mit Allium ursinum (Cardanimo-Fagetum allietosum ).
a) Struktur 1 (S1)
Diese Struktur lag im Bereich des Schönbodens und setzte
sich aus einem System von drei nahe beieinanderliegenden Baumstämmen
von ungefähr gleichem Alter zusammen. Die Baumarten waren
eine Esche, eine Buche und eine Fichte.
Die verschiedenen Waldgesellschaften waren an diesem Standort
eng verzahnt, die Strukturen lagen zu grossen Teilen in einem
Zahnwurz-Buchenwald mit Allium ursinum (Cardanimo-Fagetum allietosum
) und in einem Waldhirse-Buchenwald mit Pulmonaria sp. (Milio-Fagetum
pulmonarietosum ).
b) Struktur 2 (S2)
Die Struktur bestand aus einer über den Kellerbodenbach geworfenen
Esche und einer Buche, welche im Laufe der Vegetationszeit wieder
austrieb. Die Waldgesellschaft ist ein typischer Waldhirsen-Buchenwald
(Milio-Fagetum typicum ). Die Bachseiten im Bereich der besammelten
Struktur waren aber dicht mit Carex pendula zugewachsen (Abb.
9).
c) Struktur 3 (S3)
S3 wurde aus einem Fichten-Asthaufen und mehreren armdicken Buchenästen
gebildet, die über dem gleichen Bach wie S2 abgelagert worden
waren. Die Vegetation war identisch mit S3.
A B B I L D U N G (in Arbeit) Abb. 9: Struktur 3 über dem Kellerbodenbach, Sihlwald bei Sihlbrugg, Mai 1994
d) Struktur 4 (S4)
Auf einer Lichtung auf dem Kellerboden lagen eine Weisstanne,
eine Esche und eine Buche, die sich teilweise im Halbschatten
befand. Ausser der Esche waren alle Strukturen natürlichen
Ursprungs. Die Weisstanne war auf ca. 3 m Höhe abgebrochen.
Die Wiese war sehr feucht und setzte sich u.a. aus Ranunculus
repens, Juncus effusus und diversen Grasarten zusammen. Die tote
Buche lag teilweise in einem typischen Waldmeister-Buchenwald
(Galio odorati-Fagetum typicum ).
e) Struktur 5 (S5)
Bei dieser Struktur handelte es sich um einen Asthaufen aus Buchen-
und Eschen-Ästen sowie aus Stammteilen einer Buche. Das Material
lag in einem typischen Waldmeister-Buchenwald (Galio odorati-Fagetum
typicum ) im Bereich des Kellerbodens und stammte von einen Forsteingriff.
f) Struktur 6 (S6)
Am Wegrand in der Nähe des Streubodens lagen zwei Buchenstämme
mit dichtem Moosbewuchs, deren Durchmesser rund 80 cm. betrug.
Die dichte Belaubung liess keine Besonnung dieses Standortes zu,
weshalb die Stämme ständig feucht waren. Am Standort
stockte ein Zahnwurz-Buchenwald mit Allium ursinum (Cardamino-Fagetum
allietosum ).
g) Struktur 7 (S7)
Diese Struktur beim sogenannten SAC-Hau war ein Asthaufen menschlichen
Ursprungs, der ausschliesslich aus Fichtenteilen und Buchenästen
bestand. Im Laufe der Vegetationszeit wurde die ganze Struktur
von Impatiens Noli-tangere überwuchert und konnte ab Juli
nicht mehr besammelt werden. Bei der Waldgesellschaft handelt
es sich um einen Waldhirsen-Buchenwald mit Pulmonaria sp, Ausbildung
mit Stachys sylvatica (Milio-Fagetum pulmonarietosum, Stachys-Variante).
In der untenstehenden Tabelle 6 sind weitere Daten zu den einzelnen Strukturen zusammengefasst. Der Einfachheit halber wurden im Feld nur Schätzdaten aufgenommen, die als Prozentzahlen in die Tabelle Eingang gefunden haben.
Tab. 6: Parameter zur Beschreibung der besammelten Totholz-Strukturen. ST = Stammteile; AM = Astmaterial; R = Reisig
Grad] | [%] | |||||
| [ | ||||||
Alle gefangenen Dipteren wurden mit Essigsäureethylester ("Essigäther") abgetötet und nach rund 10 min. auf Minutien oder feinen Insektennadeln aufgespiesst. Die meisten Tiere wurden leicht seitlich durch den Thorax gestochen, Drosophilidae, Dolichopodidae sowie kleine Empididae auch seitlich durch die Pleuren, damit wichtige Kennzeichen an der Beborstung des Mesonotums nicht zerstört wurden.
Die Bestimmung der Tiere erfolgte mit Hilfe eines Wild Binokulars M5 bei 6 bis 50-facher Vergrösserung. Die umfangreiche Bestimmungsliteratur ist in Kapitel 7.2. aufgelistet. Äussere Merkmale genügten bei den meisten Arten zur Bestimmung. Bei einigen schwierig zu bestimmenden Gruppen wurden Genitalpräparate angefertigt. Dazu wurde das Abdomen möglichst vollständig abgetrennt und 24 h lang in 10%ige KOH-Lösung eingelegt oder 1 h im Trockenschrank auf 60¡C erhitzt. Danach wurden die Präparate eine halbe Stunde in Eisessig, danach zur Entwässerung in Isopropanol gegeben. Die anschliessende Präparation der Genitalien erfolgte in Glycerin mit Hilfe von feinen Nadeln. Die Genitalteile wurden nach der Bestimmung in Glycerin eingelegt.
Weibchen konnten teilweise nicht bis zur Art angesprochen werden. Die Syrphidae, Stratiomyidae [pp.], Ephydridae und Pipunculidae wurden von B. Merz, die Drosophilidae und Aulacigastridae von Dr. G. Bächli, Universität Zürich, bestimmt.
Um die Ähnlichkeiten zwischen den einzelnen Strünken zu berechnen, wurde eine Cluster-Analyse und eine Ordination durchgeführt. Eine geeignete Einführung und Zusammenfassung zu diesen komplexen Vorgehen, über die Berechnung und Bedeutung von verschiedenen Ähnlichkeitsindices und die Transformation der Werte für die Auswertung ökologischer Daten finden sich in KREBS (1987) und WILDI (1986).
Die Dendrogramme wurden mit dem Korrelationskoeffizienten ohne Transformation der Daten (weder skalar noch vektoriell) errechnet und mit der Methode "nearest neighbor (single linkage)" (KREBS, 1987) mit Hilfe des Softwareprogramms MULVA-5 (M5-LC) Version 1.0c erstellt.
Die Ordination wurde mit dem gleichen Programm durchgeführt. Die Daten wurden nicht transformiert und einer Hauptkoordinatenanalyse (PCA, coeff. normalized) unterzogen, um die in Kap. 4.2.5. gezeigte Abbildung zu erhalten.
Für die Darstellung der Phänologie wurde die Sammelperiode, während der die Fallen betrieben wurden, in Pentaden eingeteilt und die Individuenzahlen in Pentaden-Summen zusammengefasst. Als Anfangspunkt für die Fünftages-Skala wurde der Zeitpunkt genommen, an dem die Fallen aufgestellt wurden. In die Auswertung wurden nur die fünf Arten aufgenommen, die in mehr als 20 Individuen gefangen werden konnten. Bei jeder Art ist der Median ausgerechnet worden und in der Graphik mit einem Pfeil und dem Datum bezeichnet.
Die Handfänge wurden nicht speziell ausgewertet, da sie mehr ein qualitatives Gutachten sind als eine quantitative Aufsammlung. Zudem wurde nur ein Teil des gesammelten Materials bestimmt. Die Resultate sind v.a. in Kap. 4.1. Tab. 7 eingeflossen.
Im Sihlwald wurden keine Klimamessungen unternommen. Sämtliche verwendeten Daten wurden an der Eidgenössischen Forschungsanstalt Wädenswil gemessen und wurden über die Schweizerische Meteorologische Anstalt, Zürich, bezogen. Aus den Tageswerten wurde ein Pentaden-Durchschnitt errechnet (vgl. Abb. 10 und Abb. 11).
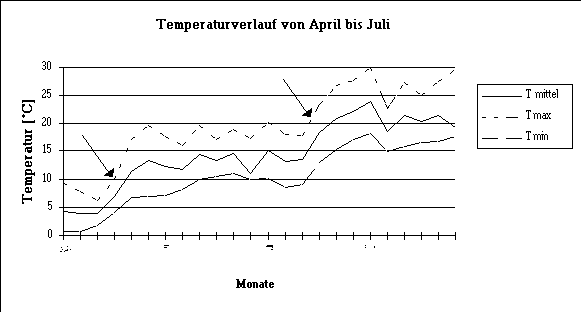
Abb. 10: Verlauf von Durchschnitts- , Maximal- und Minimaltemperaturen während der Untersuchungsperiode bei Wädenswil. Die Pfeile bezeichnen markante Temperaturanstiege.
Die Verläufe für Mittel-, Maximal- und Minimaltemperaturen sind einander ähnlich. Markante Temperaturanstiege konnten Mitte April und Mitte Juni verzeichnet werden. Dazwischen lag eine Phase mit relativ viel Niederschlag und geringerer Sonneneinstrahlung. Von Mitte Juni bis Anfang Juli stieg die Mitteltemperatur um 10¡C an und erreichte am 4.7. mit 25,4¡C das Maximum. Das Temperatur-Minimun kam auf den 15. April mit 3,4¡C zu liegen. In dieser Pentade erreichte auch die Sonnenscheindauer einen Tiefstwert mit durchschnittlich 20 min./Tag. Am meisten Niederschlag wurde in der 4. Mai-Pentade ermittelt, die Fünftages-Summe erreichte 70,5 mm.

Abb. 11: Niederschlag und Sonnenscheindauer während der Untersuchungsperiode bei Wädenswil. Für die Einstrahlung wurde ein Fünftagesmittel aufgezeichnet, für die Niederschlagsintensität die Pentadensumme.